Indukowanie mutacji Konopi
Czyli jak pomagać ewolucji
Zaawansowani ogrodnicy często są znudzeni uprawą ogólnie dostępnych na rynku odmian konopi. Jeżeli mają jeszcze możliwość swobodnej i bezpiecznej uprawy nic nie stoi, żeby spróbować pomóc ewolucji indukować mutacje, z których być może wyrosną bardzo interesujące okazy. W tym artykule przyjrzymy się czynnikom mutagennym powodującym zmianie w genotypie roślin.
Mutacje roślin
Termin po raz pierwszy wprowadzony przez biologa Hugo de Vriesa na przełomie XIX i XX wieku, oznaczający zmianę zapisu informacji genetycznej, pierwotne źródło zmienności genetycznej organizmów. Mutacja jest zjawiskiem losowym, podlegającym jednak wpływom środowiska (mutagenom – np. chemicznym, promieniowaniu). Częstość mutacji nie jest stała pomiędzy gatunkami i zależy między innymi od doskonałości aparatu powielania DNA i jego naprawy.
Sadzonka z wyraźnie zmienionymi liśćmi.
dmienną klasą mutacji są zmiany spowodowane transpozycją (tzw. skaczące geny), gdzie odcinek DNA o długości kilkuset do kilku tysięcy nukleotydów zmienia położenie w obrębie genomu. Tego typu mutacje są bardziej rozpowszechnione u niektórych roślin np. kukurydza. Wprowadzanie mutacji do materiału genetycznego nazywa się mutagenezą. Współczesna genetyka umożliwia mutagenezę ukierunkowaną, realizowaną za pomocą inżynierii genetycznej.
Celem doświadczalnych modyfikacji genomów roślinnych jest zrozumienie fundamentalnych procesów biologicznych. W przypadku roślin kwestie o podstawowym znaczeniu to proces różnicowania tkanek oraz mechanizm indukowania ekspresji poszczególnych genów przez światło. Inni badacze dążą do ulepszenia roślin ważnych dla rolnictwa. Nowe i zmienione geny wprowadza się za pomocą specjalnych wektorów do pojedynczych komórek roślinnych hodowanych w laboratorium. Z poszczególnych komórek można uzyskać całą roślinę, nawet jeśli nie są to komórki zarodkowe.
W pełni zmutowany krzak w początkowej fazie kwitnienia.
W wielu eksperymentach prowadzonych na roślinach wykorzystuje się specjalne wektory pochodzące z plazmidów bakteryjnych. W naturze występują one w komórkach bakterii powodującej raka szyjki korzeniowej bardzo wielu roślin – Agrobacterium tumefaciens. Plazmidy wraz z komórkami bakteryjnymi stanowią naturalny mechanizm wprowadzania genów do roślin. Komórki A. tumefaciens wnikają do zranionych części roślin. Plazmid przedostaje się z bakterii do komórek roślinnych i część plazmidowego DNA zostaje wbudowana do genomu gospodarza. Niektóre geny plazmidowego DNA ulegają ekspresji w komórkach roślinnych. W efekcie rozwija się guz. Co dziwne, z takich rakowatych komórek można otrzymać całe, zdrowe rośliny. Jest to jedyny znany przykład naturalnego przekazywania genów komórkom eukariotycznym przez plazmidy bakteryjne. System ten wykorzystano w inżynierii genetycznej roślin – obce geny, wstawione do plazmidu A. tumefaciens za pomocą technik rekombinacji DNA, wprowadza się następnie do roślin.
Z tak transformowanych komórek otrzymano rośliny zdolne do wytwarzania zdrowych nasion, przenoszących funkcjonalne obce geny. Geny wzmacniające odporność roślin na choroby wirusowe lub herbicydy oraz kodujące związki owadobójcze zostały już wprowadzone do roślin ważnych dla rolnictwa. Wiele z nich przechodzi obecnie próby polowe. Inne badania koncentrują się na poprawieniu wartości odżywczej roślin uprawnych.
Pierwsze oznaki mutacji.
Rodzaje mutacji:
-
genowe (punktowe) - zachodzą na odcinku DNA krótszym niż jeden gen; polegają na zmianie właściwej sekwencji nukleotydów (zamianie, wycięciu lub wstawieniu par pojedynczych nukleotydów lub odcinków trochę dłuższych)
-
chromosomowe - dotyczą zmiany struktury chromosomów lub ich liczby
-
strukturalne (aberracje) - polegają na zmianie struktury w obrębie jednego chromosomu lub pomiędzy chromosomami niehomologicznymi
-
liczbowe (genomowe) - dotyczą zmiany całego genomu, który zostaje zubożony lub powiększony o jeden chromosom lub też zwielokrotniony całkowicie (o całe "n"); są wynikiem zaburzenia procesów podziałowych, konkretnie nieprawidłowego rozejścia się chromosomów
Na tym liściu wyrósł zmutowany kwiat.
Czynniki mutagenne:
Czynniki mutagenne wywołują zmiany w materiale genetycznym roślin – powodują tak zwana mutagenezę. Mutageneza jest wykorzystywana w hodowli roślin uprawnych w celu wyprowadzenia nowych odmian. Jest też wykorzystywana w badaniach naukowych (np. analiza ekspresji genów). Mutageneza może być losowa lub ukierunkowana – wywołująca mutację w określonym miejscu w sekwencji DNA. Skutki mutacji mogą być w ogóle niezauważalne, mogą prowadzić do nieznacznej i niegroźnej zmiany fenotypu, a mogą również być przyczyna poważnych zaburzeń oraz śmierci.
Jak już wspomniano, mutacje mogą zachodzić spontanicznie oraz pod wpływem różnych czynników zewnętrznych. Takimi czynnikami mutagennymi są:
Mutageny chemiczne:
- niektóre kwasy (np. kwas azotawy)
- aminy (np. anilina)
- pestycydy
- niektóre gazy bojowe (iperyt, jako czynnik alkilujący DNA)
- barwniki wiążące DNA (interkalujące)
- dioksyny
- niemal wszystkie związki aromatyczne
- kwas azotowy (III) - HNO2 - powoduje usunięcie grup aminowych z zasad azotowych, co powoduje np. zamianę cytozyny w uracyl
- związki alkilujące (np. iperyt i jego pochodne) - powodują dołączanie do zasad azotowych grup alkilowych, co również zmienia ich charakter
- analogi zasad azotowych (np. bromouracyl) - nie są prawidłowo odczytywane podczas transkrypcji
- barwniki akrydynowe (np. oranż akrylowy, akryflawina, proflawina) - powodują wstawianie lub wycinanie sekwencji nukleotydowych
- alkaloidy - np. kolchicyna, blokująca tworzenie wrzeciona podziałowego, co powoduje, że chromosomy nie rozchodzą się podczas podziału
- sole metali ciężkich
Zbliżenie na kwiat na liściu.
Mutageny fizyczne:
- promieniowanie jonizujące
- promieniowanie gamma
- promieniowanie rentgenowskie (promieniowanie X)
- promieniowanie ultrafioletowe
- promieniowanie kosmiczne
- wysoka temperatura
- szok termiczny
- bodźce traumatyczne - urazy
Mutageny biologiczne:
- niektóre wirusy np. retrowirusy
- grzyby pleśniowe
Mutageny metaboliczne:
- brak jonów Mg2+
- brak jonów Ca2+
Najpoważniejszymi w skutkach są mutacje genomowe, czyli takie, które powodują zmianę liczby chromosomów w genomie. W wielu przypadkach są one śmiertelne już na etapie zygoty. Prawidłowy genom komórek autosomalnych zawiera 2n chromosomów, komórki rozrodcze natomiast mają 1n.
Zwielokrotnione „palce” liści.
Rośliny poliploidalne
Rośliny poliploidalne mają sztucznie zwielokrotniony cały garnitur chromosomów np. 2n, 3n, 4n …itd. Mutacje tę można wywoływać sztucznie np. kolchicyną. Zatrzymuje ona podział komórki na etapie metafazy poprzez uniemożliwienie wytworzenia mikrotubul wrzeciona kariokinetycznego, co uniemożliwia rozejście się chromosomów.
U roślin poliploidalnych ma miejsce powiększenie objętości komórek, co powoduje gigantyzm organów; nie wszystkie organy jednak wykazują takie samo powiększenie. Występuje także mniejsza liczba aparatów szparkowych, co zmniejsza powierzchnię transpiracji, a to z kolei czyni rośliny bardziej odpornymi na suszę. Najbardziej korzystne do uprawy są rośliny tetraploidalne (4n), większa liczba chromosomów powoduje już zaburzenia w wykształcaniu organów.
Większość roślin tego typu nie tworzy płodnych nasion, więc jest mała szansa na przeniesienie mutacji na przyszłe pokolenia. Należy zaznaczyć, iż tylko parzyste poliploidy są płodne (na drodze mejotycznego podziału nieparzysta liczba chromosomów nie rozdzieli się po równo). Dlatego komórki poliploidalne muszą zachowywać się jak komórki diploidalne, aby ich chromosomy były w stanie wytwarzać homologiczne pary.
Wg różnych szacunków od 35%[1] do 47%[2] wszystkich roślin kwiatowych ma liczbę chromosomów będącą wielokrotnością najniższej liczby występującej w danym rodzaju. Ok. 75% roślin naczyniowych odziedziczyło najprawdopodobniej zwielokrotnione zestawy chromosomów od dawnego, poliploidalnego przodka[1]. We florze środkowej Europy mniej więcej połowa roślin jest naturalnymi lub wytworzonymi przez człowieka poliploidami (można tego dokonać np. poprzez zastosowanie kolchicyny, która niszczy wrzeciono kariokinetyczne uniemożliwiając podział komórki). Jako przykłady można podać pszenżyto, ziemniaki, wiele roślin ozdobnych i traw, drzewa owocowe. Odznaczają się one większymi rozmiarami, większą żywotnością i odpornością (dają, więc także większe plony).
Jednak w przypadku roślin możliwość krzyżowania jest bardziej skomplikowana. Nowy poliploid może dawać płodne potomstwo tylko wtedy, gdy jego gamety połączą się z gametami o tym samym stopniu ploidalności. Dlatego też poliploidyzacja jest u roślin ważnym mechanizmem specjacji. Na przykład, jeśli tetraploid wytworzy gamety o liczbie chromosomów 2x i połączą się one z gametami haploidalnymi x, potomstwo będzie triploidem (3x, a więc będzie bezpłodne). W takim wypadku płodność może odzyskać jedynie, gdy powstaną w nim komórki heksaploidalne (na skutek nieprawidłowej mitozy), które to komórki będą w stanie tworzyć żywotne gamety. Wiele roślin jest zdolnych do rozmnażania wegetatywnego, które pozwala im "przeczekać" do czasu przywrócenia płodności. Rośliny mają także przewagę ze względu na brak komórek szlaku płciowego (z każdej komórki zdolnej do podziału może powstać klon, u którego w gametoficie może dojść do podwojenia ilości chromosomów i odzyskania płodności).
UWAGA! Nie należy mylić poliploidalności z aneuploidalnością, która charakteryzuje się niewłaściwą liczbą pojedynczych chromosomów.
Poliploidy bardzo często powiększają komórki (zwłaszcza u roślin, co prowadzi do gigantyzmu, u zwierząt zwykle się go nie obserwuje, co jest prawdopodobnie zasługą sprawniejszych mechanizmów regulacji rozwoju - większe komórki są rekompensowane ich mniejszą liczbą). Poliploidy dojrzewają wolniej w porównaniu z osobnikami diploidalnymi. Poliploidalność często jest wykorzystywana dla zwiększania atrakcyjności produktów (np. większe komórki poprawiają strukturę owoców i warzyw takich jak truskawki, jabłka czy ziemniaki). Sterylność triploidów jest przydatna przy produkcji owoców beznasiennych (np. beznasienne ogórki są mniej gorzkie). Poliploidyzacja prowadzi do stabilizacji genomów (może eliminować skrajne genotypy).
Zaawansowane techniki uprawy konopi wymagają sporej wiedzy i dużo szczęścia. Jednak ich uprawa przynosi bardzo wiele radości i satysfakcji, a rezultaty mogą zadziwić nawet profesjonalnych producentów nasion z firm komercyjnych. Należy przygotować się jednak na sporo porażek i dziwnych deformacji. Najczęstsze są zmiany w budowie liści i kwiatów. Zmienia się także wydajność roślin i czas ich dojrzewania.
Rekombinacja/sprzężenie
Diploidalna natura chromosomów pozwala genom innych chromosomów na dobór niezależny podczas rozmnażania płciowego rekombinując nową kombinację genów. Geny tego samego chromosomu teoretycznie nigdy nie będą rekombinowane. Podczas krzyżowania się, chromosomy wymieniają odcinki DNA efektywnie mieszając allele genowe między chromosomami. Proces krzyżowania chromosomów następuje podczas mejozy – serii podziału, w wyniku której powstają komórki haploidalne.
Prawdopodobieństwo krzyżowania się chromosomów następującego między dwoma punktami genów odnosi się do odległości między dwoma punktami. Dla dowolnie długiego dystansu prawdopodobieństwo krzyżowania się jest wystarczająco wysokie, aby dziedziczenie genów nie korelowało ze sobą – allele dwóch genów dążą, aby być dziedziczone wspólnie. Liczbę krzyżowań między seriami genów można ustalić na podst. linearnej mapy krzyżowań, która ogólnie opisuje układ genów wzdłuż chromosomów.
Pierwsze prawo Mendla.
Nazywane jest prawem czystości gamet oraz segregacji. Prawo to zakłada że:
- Każda cecha dziedziczna jest określona przez jednostki dziedziczności (tę nazwę wprowadził Mendel, dzisiaj nazywane są genami) występujące parami.
Jeśli obydwa geny określające daną cechę są identyczne, wówczas organizm taki nazywamy homozygotą.
Jeśli geny określające daną cechę są różne, wówczas taki organizm nazywamy heterozygotą (mieszańcem). Przeciwstawne formy tego samego genu nazywane są allelami. Sformułowanie Mendla można zilustrować za pomocą wygodnego, powszechnie stosownego systemu znaczeń literowych. W zapisie dużą literą oznaczamy gen dominujący, a taką samą literą tylko małą odpowiadający mu gen recesywny.
Zapis symboliczny osobnika homozygotycznego
AA – homozygota dominująca
aa – homozygota recesywna
Zapis symboliczny osobnika heterozygotycznego
Aa – heterozygota
- W czasie tworzenia się gamet każda para genów podlega segregacji (rozdzieleniu) w taki sposób, że każda gameta otrzymuje tylko jeden gen z danej pary (I prawo Mendla). To oznacza, że gameta zawiera jeden rodzaj genu warunkującego daną cechę.
Pierwsze prawo odnosi się do dziedziczenia jednej cechy i można je przedstawić graficznie (przy pomocy krzyżówki).
Krzyżówka – czerwono kwitnącego grochu z grochem kwitnącym biało – nazywana krzyżówką – typ pisum (dominacji zupełnej).
Dane:
A– oznacza gen zapisujący barwę czerwoną,
a– oznacza gen zapisujący barwę białą,
P– pokolenie rodziców,
F1, F2 – kolejne pokolenia
Skrzyżowano osobnika o czerwonych kwiatach z osobnikiem o białych kwiatach
P AA x aa
Gamety A, A, a, a
F1 Aa Aa Aa Aa
- Można ustalić cechy poszczególnych roślin:
Fenotyp, czyli zewnętrzny wygląd osobnika będący efektem współdziałania jego genów i warunków środowiska.
W uzyskanym pokoleniu F1 wszystkie osobniki będą miały czerwone kwiaty (Aa).
Genotyp to zespół genów danego osobnika. W otrzymanym pokoleniu F1 wszystkie osobniki będą heterozygotami.
W kolejnym pokoleniu po skrzyżowaniu osobników z pokolenia F1 otrzymamy.
Aa x Aa
F2 AA Aa Aa aa
czyli:
fenotyp będzie 3:1 (3 czerwone i 1 biały)
genotyp będzie 2:2 (2 homozygoty /AA, aa/, 2 heterozygoty /Aa/).
Aby stwierdzić, czy osobniki wykorzystywane do doświadczenia (krzyżówki) są homozygotami dominującymi czy heterozygotami (fenotyp będzie identyczny) przeprowadzić należy krzyżówkę testową – polegającą na skrzyżowaniu osobnika z homozygotą recesywną. Jeśli badana roślina była homozygotą dominującą wówczas wszystkie rośliny w F1 będą kwitły na czerwono. Natomiast jeśli połowa roślin F1 będzie miała białe kwiaty to badana roślina była heterozygotą. Rozkład fenotypów w krzyżówce testowej wynosi 2:2.
W wieloletnich pracach, zwłaszcza przy zmianie obiektów doświadczenia, Mendel zauważał inne sposoby przekazywania i ujawniania się cech u osobników potomnych. Z taką innością spotkał się Mendel, kiedy skrzyżował rośliny dziwaczka o kwiatach białych i czerwonych. Okazało się, że w pierwszym pokoleniu powstała fenotypowa cecha będąca połączeniem cech rodziców. U tych roślin allel determinujący barwę kwiatów wykazuje niepełną dominację, czyli allele kodują odmienne cechy, lecz żaden z nich nie dominuje nad drugim (współdominowanie).Powstanie cechy pośredniej można wytłumaczyć zbyt małą ilością wyprodukowanego barwnika (przy obecności jednego allelu dominującego), który nie wystarcza do pełnego zabarwienia kwiatów tworząc fenotyp pośredni.
Krzyżówka z niepełną dominacją – typ zea
P AA x aa (białe)
F1 Aa x Aa (różowe)
Aa x Aa (różowe)
F2 AA Aa Aa aa
czerwone różowe różowe białe
stosunek fenotypów wynosi 1 : 2 : 1
Genotyp – jeśli allele wykazują niepełną dominację trudno powiedzieć, który z alleli można uznać za dominujący.
Drugie prawo Mendla
Dotyczy krzyżówek, w których rodzice różnią się większą liczbą niezależnych cech.
Każda para genów podlega losowej segregacji i jest dziedziczona niezależnie od genów innych.
Obecnie wiadomo, że prawo niezależnej segregacji sprawdza się tylko wtedy, gdy każda para genów znajduje się na innej parze homologicznych chromosomów. Gdy są na jednym chromosomie, będą przekazywane razem, ponieważ to chromosomy ulegają niezależnej segregacji, a nie geny jak uważał Mendel.
Krzyżówka przedstawiająca drugie prawo Mendla
Mendel przeprowadził doświadczenie, w którym badał 2 cechy morfologiczne, za które odpowiedzialne są różne geny dziedziczące się niezależnie od siebie.
Obserwował barwę i pokrycie nasion grochu.
Dane:
A, a – barwa nasiona (A-żółte; a-zielone)
B, b – kształt nasion (B-gładkie; b-pomarszczone)
Skrzyżował rośliny o żółtych, gładkich nasionach z zielonymi, pomarszczonymi.
P AABB x aabb
F1 Aa Bb Aa Bb Aa Bb Aa Bb
W pokoleniu F1
fenotypy były 4:0 – wszystkie potomne rośliny wytwarzały żółte gładkie nasiona
genotypy – wszystkie osobniki były heterozygotami
Aa Bb x Aa Bb
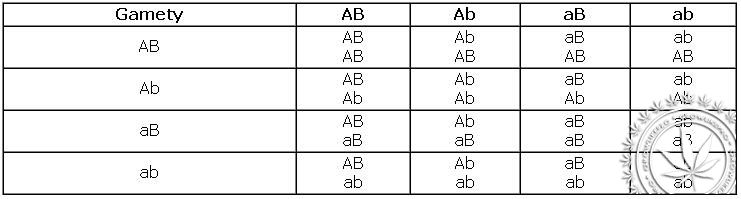
Wynik losowego połączenia się wszystkich gamet może przedstawić pod postacią szachownicy Punneta:
Gamety AB Ab aB ab
AB AB
AB Ab
AB aB
AB ab
AB
Ab AB
Ab Ab
Ab aB
Ab ab
Ab
aB AB
aB Ab
aB aB
aB ab
aB
ab AB
ab Ab
ab aB
ab ab
ab
Genotyp – 2 homo : 14 hetero
Fenotyp:
9 roślin o żółtych gładkich nasionach
3 rośliny o żółtych pomarszczonych
3 rośliny o zielonych gładkich
1 roślina o zielonych pomarszczonych
Odstępstwa od praw Mendla
Allele wielokrotne
Zgodnie z I prawem Mendla istnieją dwa allele tego samego genu. Okazało się, że jest to prawdziwe dla jednego osobnika. Są przypadki, że w całej populacji może występować więcej odmian tego samego genu. Są to allele wielokrotne. Oznacza to występowanie w populacji wielu alleli danej cechy. Jest to prawdopodobnie efektem mutacji i przyczynia się do wzrostu zmienności cech w obrębie populacji.
Przykładem alleli wielokrotnych są grupy krwi w populacji człowieka. Wyróżniamy grupy A, B, AB, O.
Osobnik posiadający określoną grupę krwi może być homo lub heterozygotą.
Grupa krwi A
IA IA zapis dla homozygoty
IA i zapis dla heterozygoty
Grupa krwi B
IB IB zapis dla homozygoty
IB i zapis dla heterozygoty
Grupa krwi AB
IA IB zapis tylko jeden
Grupa krwi O
i i zapis tylko jeden
Plejotropizm.
Według Mendla każdy gen warunkował jedną cechę organizmu. Dzięki późniejszym badaniom, udowodniono istnienie sytuacji, że jeden gen może wpływać na różne cechy organizmu. Nazwano to zjawisko plejotropowym działaniem genu. W trakcie przeprowadzanego doświadczenia na roślinach stwierdzono, że gen A wpływa na barwę płatków oraz pojawiające się plamy na łodydze.
Współdziałanie genów
Istnieje sytuacja, że ujawnienie się jednej cechy zależy od wielu genów. Doświadczalnie udowodnili takie dziedziczenie Bateson i Punnet.
Skrzyżowali oni biało kwitnący groszek pachnący ze sobą. W pokoleniu F1 otrzymali wszystkie potomne rośliny o kwiatach czerwonych. W pokoleniu F2 rozkład cech fenotypowych wynosił 9:7. Okazało się, że czerwona barwa kwiatów wywołana jest barwnikiem antocyjanem, którego powstawanie zapisują geny dominujące A, B. Osobniki, u których jeden z genów jest recesywny, mają kwiaty białe,
czyli P aa BB x AAbb
kwiat biały kwiat biały
F1 AaBb, AaBb – wszystkie kwiaty będą czerwone.
Dzisiaj wiadomo, że do cech zapisanych przez wiele genów należą u człowieka: wzrost, kształt ciała, barwa skóry, inteligencja.
Geny uzupełniające się.
Istnieją geny hipostazy oraz epistazy. Te pojęcia (epistaza, hipostaza) odnoszą się do genów, które nie są allelami. Obecność dominującego genu epistazy umożliwia ujawnienie cech zapisanych w genach hipostazy.
Geny supresorowe
Są to geny, które swoją obecnością powodują znoszenie efektów fenotypowych innych genów.
Geny letalne
Są to najczęściej geny recesywne, których obecność prowadzi do śmierci organizmu. Powodują one umieranie organizmu już w okresie rozwoju zarodkowego.
Udanych eksperymentów!
źródło: cannabis.info






")
")
")

")

")








")
")



")
")
")





