Cannabis sativa od dawien dawna stosowana byŇāa w celach leczniczych [1]. Gatunek ten zawiera kanabinoidy, czyli wyjńÖtkowńÖ grupńô zwińÖzk√≥w terpenofenolowych, kt√≥re gromadzone sńÖ w trichomach na powierzchni roŇõliny[2]. Ponad 70 r√≥Ňľnych kanabinoid√≥w zostaŇāo wyizolowanych z konopi, a najbardziej aktywnym biologicznie zwińÖzkiem okazaŇā sińô őĒ9-tetrahydrokannabinol zwany r√≥wnieŇľ őĒ9-THC [3]. Wzory 6 gŇā√≥wnych kanabinoid√≥w zostaŇāy przedstawione na rysunku 1.
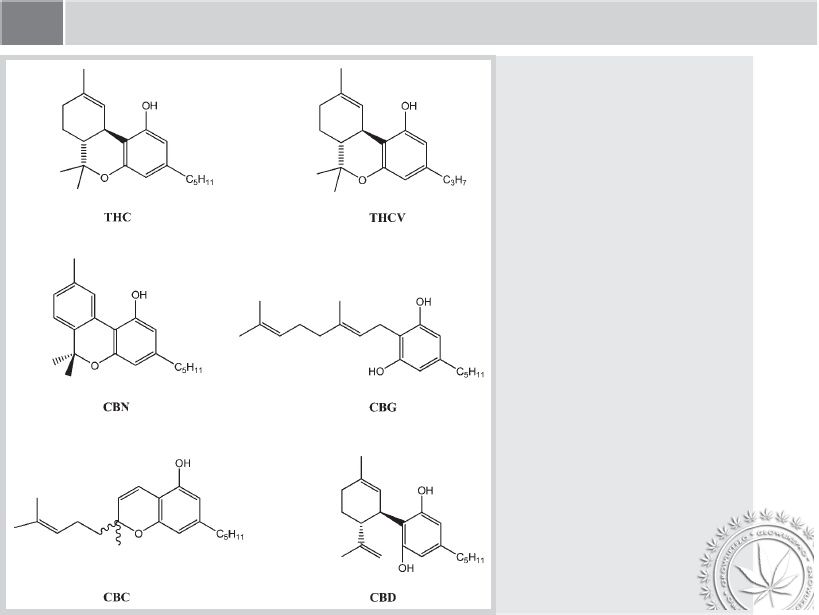
őĒ9-THC opr√≥cz swoich psychoaktywnych wŇāaŇõciwoŇõci moŇľe byńá stosowany r√≥wnieŇľ jako Ňõrodek przeciwb√≥lowy, przeciwzapalny, przeciw wymiotny lub pobudzajńÖcy apetyt, co czyni ten zwińÖzek przydatnym w celach terapeutycznych [4]. Terapeutyczna skutecznoŇõńá konopii i őĒ9-THC zostaŇāa doŇõńá obszernie opisana [5] [6] [7] [8] [9] [10]. őĒ9-THC jest aktualnie dostńôpne na rynku w postaci Ňľelatynowych kapsuŇāek do uŇľytku wewnńôtrznego, w preparacie o nazwie Marinol. Dostawa hurtowych iloŇõci syntetycznego őĒ9-THC zostaŇāa ograniczona do produkcji tylko tego preparatu, co utrudniŇāo opracowanie nowych preparat√≥w, np. czopk√≥w zawierajńÖcych őĒ9-THC. Z tego powodu rozwaŇľane jest pozyskiwanie őĒ9-THC dzińôki izolacji z materiaŇāu roŇõlinnego. KluczowńÖ sprawńÖ jest pozyskanie kwiat√≥w odpowiedniej jakoŇõci, co umoŇľliwiŇāoby uzyskiwanie őĒ9-THC przy uŇľyciu niŇľszego nakŇāadu finansowego, niŇľ w przypadku chemicznej syntezy őĒ9-THC. Wsp√≥Ňāczesne badania majńÖ na celu ulepszenie poszczeg√≥lnych odmian konopi w celu osińÖgnińôcia duŇľego plonu o wysokiej zawartoŇõci THC, kt√≥ry m√≥gŇāby byńá zastosowany do izolacji naturalnego THC do zastosowaŇĄ farmaceutycznych.
Od lat 80 zaobserwowano zwińôkszajńÖcńÖ sińô czńôstotliwoŇõńá przypadk√≥w konfiskaty marihuany w USA [11] [12]. Z powodu zapylenia krzyŇľowego jako sposobu rozmnaŇľania sińô konopi, niezwykle trudne jest utrzymanie staŇāego wysokiego poziomu produkcji THC w kwiatach, w przypadku uprawy z nasion, w warunkach polowych lub szklarniowych. Z tego powodu uŇľycie zaawansowanych technik hodowli tkankowych (mikrorozmnaŇľanie in vitro) umoŇľliwia szybkie rozmnoŇľenie wybranych roŇõlin cechujńÖcych sińô poŇľńÖdanymi przez nas parametrami. Dotychczas opracowano wiele sposob√≥w mikrorozmnaŇľania in vitro dla r√≥Ňľnych genotyp√≥w konopi i miejsc pozyskiwania materiaŇāu do klonowania [13][14][15][16][17][18], a takŇľe zaobserwowano znaczne r√≥Ňľnice w rozwoju kultur w przypadku poszczeg√≥lnych metod. PoniewaŇľ niezwykle istotne jest uzyskiwanie materiaŇāu roŇõlinnego wysokiej jakoŇõci o staŇāych parametrach, zostaŇāa opracowana metoda rozmnaŇľania in vitro przy uŇľyciu segment√≥w wńôzŇāowych Ňāodyg, zawierajńÖcych zawińÖzki bocznych pńôd√≥w.W celu sprawdzenia przydatnoŇõci tej metody por√≥wnano jńÖ z klasycznńÖ metodńÖ klonowania, pobierajńÖc klony z tej samej roŇõliny-matki w tym samym okresie wzrostu. Por√≥wnanie to polegaŇāo na chemicznej analizie zawartoŇõci kanabinoid√≥w przy uŇľyciu chromatografii gazowej.
4. MateriaŇāy i metody
RoŇõliny uŇľyte do badaŇĄ
RoŇõliny Canabis sativa uŇľyte do badaŇĄ wyrosŇāy z wysokojakoŇõciowych meksykaŇĄskich nasion w konopnym ogrodzie na uniwersytecie Misisipi w 2006 r. ZaŇāoŇľono pole skŇāadajńÖce sińô z okoŇāo 31 000 roŇõlin, a kaŇľda roŇõlina zostaŇāa otagowana przy pomocy kodu kreskowego w celu stworzenia dokŇāadnego spisu roŇõlin oraz umoŇľliwienia identyfikacji w toku dalszych badaŇĄ. Podczas kwitnienia mńôskie roŇõliny zostaŇāy usunińôte w celu uniknińôcia zapylenia. SpoŇõr√≥d pozostaŇāych ŇľeŇĄskich roŇõlin wybrano 50 zdrowych osobnik√≥w z r√≥Ňľnych poletek. MateriaŇā z tych roŇõlin analizowano pod kńÖtem zawartoŇõci kanabinoid√≥w, w r√≥Ňľnych stadiach wzrostu. W oparciu o wyniki uzyskane podczas wzrostu wegetatywnego, z kilku wybranych osobnik√≥w pobrano sadzonki w celu dalszej uprawy. Klony te przeniesiono do pomieszczenia i trzymano w fazie wegetatywnej pod kombinacjńÖ 1000 watowych lamp MH i HPS. Klony uprawiane w pomieszczeniu identyfikowano na podstawie zawartoŇõci THC i innych kanabinoid√≥w, i powińÖzano z odpowiednińÖ roŇõlinńÖ‚ÄďmatkńÖ uprawianńÖ na polu. RoŇõliny zostaŇāy dokŇāadnie oznaczone i podzielone na kategorie bazujńÖc na informacjach o ich polnych roŇõlinach-matkach. Tak wyhodowane roŇõliny posŇāuŇľyŇāy jako roŇõliny-matki uprawiane w pomieszczeniu (MP-indoor). Dla naszych badaŇĄ zostaŇā wybrany klon z roŇõliny z pola, o identyfikatorze P1-2714.
Po 6 miesińÖcach aklimatyzacji i wzrostu w fazie wegetatywnej pod oŇõwietleniem 18/6, pobrano odcinki wńôzŇāowe z wybranej roŇõliny macierzystej do mikrorozmnaŇľania in vitro (IVP). Po 6 tygodniach ustabilizowane i dobrze rozwinińôte pńôdy przesadzono do medium ukorzeniajńÖcego. W tym samym czasie,z tej samej roŇõliny-matki pobrano klasyczne klony(VP), w postaci 6-10 cm fragment√≥w Ňāodyg, i ukorzeniano w doniczkach torfowych o Ňõrednicy 5cm. Po 5 tygodniach dobrze ukorzenione IVP oraz VP przesadzono do identycznych doniczek (10cm Ňõrednicy) z identycznym medium (50% wŇā√≥kno kokosowe/50% komercyjna mieszanka glebowa). Oba rodzaje klon√≥w utrzymywano w identycznych warunkach: 25 stopni (+/-3), wilgotnoŇõńá 55% (+/-5), 7x 1000w HID, 7x 1000w HPS, 32,5 m2. Po 4 tygodniach wzrostu roŇõliny przesadzono do doniczek o Ňõrednicy 30 cm.
Po kolejnych 6 tygodniach wzrostu pobrano pr√≥bki ze szczytowych czńôŇõci roŇõlin w celu oznaczenia zawartoŇõci kanabinoid√≥w w szczycie fazy wegetatywnej. Nastńôpnie oba rodzaje klon√≥w przeŇāńÖczono na kwitnienie (12/12). ZapoczńÖtkowanie kwitnienia nastńÖpiŇāo w przecińÖgu 15 dni. Okresowe pobieranie pr√≥bek do oznaczania kanabinoid√≥w odbywaŇāo sińô w tym samym czasie we wszystkich grupach roŇõlin (klony in vitro, klony normalne, roŇõliny matki). Analizowano 9 klon√≥w IVP i 9 klon√≥w VP. Do por√≥wnania uŇľyto danych pŇāynńÖcych z analizy pr√≥bek pobranych w 15, 24,26, i 28 tygodniu Ňľycia roŇõlin. Wiek liczony byŇā od dnia pobrania klon√≥w w przypadku VP, i od dnia przeniesienia do medium ukorzeniajńÖcego w przypadku IVP.

Szczytowe segmenty wńôzŇāowe zawierajńÖce boczne pńôdy ( ok. 1 cm dŇāugoŇõci) zostaŇāy przeszczepione w celu uzyskania kultur in vitro. Przeszczepiany materiaŇā pobierany byŇā z roŇõlin-matek rosnńÖcych w pomieszczeniu (MP-indoor). Powierzchnia przeszczepianego materiaŇāu zostaŇāa zdezynfekowana przez zanurzenie w roztworze zawierajńÖcym 0,5 % NaOCl (15 % roztw√≥r wybielacza) oraz 0,1% Tween 20 przez 20 minut. Nastńôpnie pobrany materiaŇā przemywano 3 krotnie destylowanńÖ wodńÖ przez 5 minut, i zaszczepiono na poŇľywce. MikrorozmnaŇľanie i hartowanie przeprowadzano wedŇāug protokoŇāu opracowanego przez Lata i in. [19]
ŇöwieŇľe fragmenty szczytowe Ňāodyg o dŇāugoŇõci 6-10 cm zawierajńÖce co najmniej 2 wńôzŇāy zostaŇāy pobrane z tej samej roŇõliny-matki co IVP. Przy uŇľyciu sterylnego ostrza wykonano cińôcie pod kńÖtem 45 stopni zaraz poniŇľej wńôzŇāa, i od razu zanurzone w wodzie destylowanej, w celu uniknińôcia tworzenia sińô pńôcherzyk√≥w powietrza w Ňāodydze. OkoŇāo 2 cm dolny kawaŇāek sadzonki zanurzano w 0,1% IBA. Podobnie jak w przypadku mikrosadzonek in vitro, klasyczne klony umieszczono w 5 cm doniczkach torfowych zawierajńÖcych wŇā√≥kno kokosowe i mix ziemi komercyjnej w stosunku 1:1. Co najmniej jeden z wńôzŇā√≥w pozostawaŇā poniŇľej poziomu gleby w celu efektywnego ukorzenienia. Tak przygotowane klony oraz klony IVP utrzymywano w identycznych warunkach pod lampami fluorescencyjnymi. Pomimo tego Ňľe proces ukorzeniania zapoczńÖtkowany zostaŇā w cińÖgu 2-3 tygodni, wińôkszoŇõńá klon√≥w utrzymywano w tych warunkach przez 6 tygodni, w celu zapewnienia lepszego wzrostu wegetatywnego.
(tu pomijam dokŇāadny opis procedury, bo kto ma dostńôp do chromatografu gazowego ten wie, a kto nie ma dostńôpu temu nie jest to potrzebne) Pr√≥bki do badaŇĄ pobierane byŇāy ze szczytowych fragment√≥w VP , IVP i MP-indoor w r√≥Ňľnych okresach wzrostu i rozwoju roŇõlin. Z materiaŇāu roŇõlinnego kaŇľdej pr√≥bki wykonano 3 powt√≥rzenia analiz. Wykorzystano procedurńô opracowanńÖ przez Ross i in. (metoda ta byŇāa wykorzystywana przez gnidy z DEA do analiz skonfiskowanej marihuany)[20].
Analizy statystyczne wykonano przy uŇľyciu programu SAS 9,1, wykorzystujńÖc jednoczynnikowńÖ analizńô wariancji ANOVA (model efekt√≥w staŇāych) oraz testy.
Na og√≥Ňā zawartoŇõńá THC roŇõnie wraz ze wzrostem roŇõŇāiny aŇľ do najwyŇľszych wartoŇõci w szczytowym okresie kwitnińôcia, nastńôpnie obserwowany jest spadek wraz z procesem starzenia sińô roŇõliny. W warunkach polowych najwyŇľsza koncentracja THC (11,53-13,54%) zostaŇāa zaobserwowana u roŇõlin pomińôdzy 90 a 105 dniem (pomińôdzy fazńÖ III i IV).Spadek zawartoŇõci THC zostaŇā zaobserwowany u roŇõlin 120-dniowych. InteresujńÖcym jest fakt, Ňľe stńôŇľenie THCV stanowiŇāo mniej niŇľ 1% caŇākowitego stńôŇľenia THC w okresie zbior√≥w. THCV bńôdńÖc homologiem THC, posiada bardzo podobnńÖ strukturńô chemicznńÖ do THC, co utrudnia rozdzielenie tych zwińÖzk√≥w. WyŇľsze stńôŇľenie THCV w materiale roŇõlinnym powoduje Ňľe proces izolacji czystego THC staje sińô droŇľszy i bardziej skomplikowany. W wyniku tego proces selekcji roŇõlin-matek na podstawie stńôŇľenia kanabinoid√≥w moŇľe mieńá kluczowńÖ rolńô w procesie izolacji THC do cel√≥w farmaceutycznych. W zwińÖzku z tym na roŇõlinńô matkńô w tym eksperymencie wybrano osobnika o wysokim stńôŇľeniu THC a niskim stńôŇľeniu THCV.
Klony z roŇõliny matki rosnńÖcej na zewnńÖtrz utrzymywano w fazie wegetatywnej w kontrolowanych warunkach przez 6 miesińôcy. Po tym okresie fragmenty Ňāodyg zawierajńÖce boczne pńôdy zostaŇāy uŇľyte do klonowania in vitro. Wybrana metoda mikrorozmnaŇľania roŇõlin jest relatywnie Ňāatwa i cechuje sińô wysokńÖ skutecznoŇõcińÖ. Ponadto obrana metoda oparta o fragmenty z bocznymi pńôdami zmniejsza ryzyko genetycznej niestabilnoŇõci roŇõliny, gdyŇľ istniejńÖce juŇľ merystemy sńÖ bardziej odporne na powstawanie zmian w genomie, w por√≥wnaniu do metod opartych na r√≥Ňľnicowaniu sińô kallusa.
Najlepszy wynik w przypadku indukcji pńôd√≥w zaobserwowano w przypadku medium Murashige‚Äôa i Skoog‚Äôa zawierajńÖcego 0,5 ¬ĶM TDZ. Dobrze rozwinińôte pńôdy zostaŇāy przeniesione do medium zawierajńÖcego 0,25 ¬ĶM TDZ, uzupeŇānionego o wńôgiel aktywowany oraz r√≥Ňľne stńôŇľenia IAA, IBA oraz NAA w celu ukorzenienia sadzonek. NaleŇľy zaznaczyńá, Ňľe klasyczne klony zostaŇā pobrane w momencie gdy klony in vitro zostaŇāy przeniesione do medium ukorzeniajńÖcego. Oba rodzaje klon√≥w utrzymywano w fazie wegetatywnej w takich samych warunkach pod lampami fluorescencyjnymi. NajwyŇľsze prawdopodobieŇĄstwo ukorzenienia w przypadku klon√≥w in vitro zaobserwowano w przypadku poŇľywki zawierajńÖcej 0,25 ¬ĶM TDZ, 500mg/l wńôgla aktywowanego, oraz 2,5 ¬ĶM IBA. Pr√≥bki do analiz stńôŇľenia kanabinoid√≥w zostaŇāy pobrane ze wszystkich 3 grup roŇõlin zaraz przed przeŇāńÖczeniem oŇõwietlenia na 12/12. Kwitnienie zostaŇāo zaobserwowane po 15 dniach w przypadku roŇõlin matek, oba rodzaje klon√≥w zakwitŇāy po 17 dniach.
Badanie to miaŇāo na celu okreŇõlenie czy klony in vitro (hodowane przy uŇľyciu specjalnej procedury[19] ) bńôdńÖ miaŇāy identyczne stńôŇľenie kanabinoid√≥w co klony pobrane w klasyczny spos√≥b. Istotne r√≥Ňľnice pomińôdzy roŇõlinami-matkami a klonami in-vitro obserwowano w przypadku wykorzystania innych metod mikrorozmnaŇľania, w wyniku zachodzenia wielu mutacji somatycznych [24] [25] [26] [27]. PoniewaŇľ celem tego badania byŇāo opracowanie metody mikrorozmnaŇľania in vitro, kt√≥ra mogŇāa by byńá uŇľyta w komercyjnych projektach uprawy Cannabis sativa, niezbńôdne byŇāo upewnienie sińô, Ňľe wybrana metoda nie powoduje nieporzńÖdanych mutacji majńÖcych wpŇāyw na metabolizm, a co za tym idzie na stńôŇľenie kanabinoid√≥w. W tym celu badano stńôŇľenia kanabinoid√≥w roŇõlin matek i pobranych klon√≥w w r√≥Ňľnych stadiach rozwoju roŇõlin.
StńôŇľenie kanabinoid√≥w w 3 badanych grupach roŇõlin (klony VP i IVP, MP-indoor) okazaŇāo sińô identyczne. Podobne wyniki otrzymali Ma i Gang [28] w przypadku imbiru uprawianego w szklarni i klon√≥w pobranych z tych roŇõlin. StabilnoŇõńá genetyczna mikrosadzonek hodowanych przy uŇľyciu opracowanej przez nas metody zostaŇāa sprawdzona przy uŇľyciu marker√≥w ISSR [29].
R√≥Ňľnice w zawartoŇõci őĒ9-THC podczas poszczeg√≥lnych faz wzrostu i rozwoju roŇõlin zostaŇāy przedstawione na rysunku 4. NajwyŇľsze stńôŇľenie őĒ9-THC zostaŇāo osińÖgnińôte w 24 tygodniu Ňľycia roŇõlin i utrzymywaŇāo sińô przez okoŇāo 2 tygodnie.
Spadek stńôŇľenia CBC wraz z wiekiem obserwowany byŇā we wszystkich grupach roŇõlin. Najsilniejsze zmniejszenie zawartoŇõci CBC wystńôpowaŇāo zaraz po zakoŇĄczeniu fazy wegetatywnej, w p√≥Ňļniejszych okresach spadek ten byŇā mniej intensywny. Zmiany stńôŇľenia CBG w czasie trwania Ňľycia roŇõlin byŇāy podobne do zmian obserwowanych w przypadku THC, THCV i CBD. We wszystkich badanych grupach roŇõlin najwyŇľsze stńôŇľenie CBG przypadaŇāo na szczyt fazy kwitnińôcia (5x wyŇľsze stńôŇľenie niŇľ w fazie wegetatywnej), a podczas okresu starzenia sińô roŇõliny iloŇõńá tego kanabinoidu zmniejszaŇāa sińô.
W przeciwieŇĄstwie do CBC, w przypadku CBN zaobserwowano staŇāy przyrost jego zawartoŇõci w cińÖgu wszystkich faz wzrostu roŇõlin, od fazy wegetatywnej do fazy zamierania. W fazie starzenia sińô roŇõliny zaobserwowano istotne statystycznie r√≥Ňľnice w stńôŇľeniu CBN mińôdzy obydwoma rodzajami klon√≥w. W okresie zbior√≥w brak istotnych r√≥Ňľnic pomińôdzy grupami. Por√≥wnanie wszystkich trzech grup roŇõlin (VP,IVP,MP-indoor) do roŇõliny matki uprawianej na dworze wykazaŇāo, Ňľe charakteryzowaŇāa sińô ona wyŇľszńÖ zawartoŇõcińÖ THC. Wszystkie badane roŇõliny byŇāy identyczne genetycznie, wińôc fakt ten moŇľna wytŇāumaczyńá r√≥Ňľnicami w warunkach wzrostu pomińôdzy uprawńÖ wewnńÖtrz i na dworze. Podczas sŇāonecznego letniego dnia w Mississippi natńôŇľenie ŇõwiatŇāa wynosi okoŇāo 1500 ¬Ķmol*m‚ąí2*s‚ąí1, podczas gdy intensywnoŇõńá ŇõwiatŇāa w pomieszczeniu wynosiŇāa okoŇāo 700 ¬Ķmol*m‚ąí2*s‚ąí1. W poprzednich badaniach zanotowano szybsze zachodzenie procesu fotosyntezy oraz lepszy wzrost w 30 stopniach C i przy natńôŇľeniu ŇõwiatŇāa 1500 ¬Ķmol*m‚ąí2*s‚ąí1.
PodsumowujńÖc, roŇõliny wyhodowane przy uŇľyciu metod in vitro nie r√≥ŇľnińÖ sińô zawartoŇõcińÖ kanabinoid√≥w od roŇõlin wyhodowanych przy pomocy tradycyjnych klon√≥w, a takŇľe od roŇõlin matek z kt√≥rych pobierano materiaŇā do klonowania. Wyniki te potwierdzajńÖ identycznoŇõńá klon√≥w in vitro i roŇõlin matek, a takŇľe sugerujńÖ Ňľe klonowanie in vitro nie wprowadza znaczńÖcych zmian w mechanizmach biochemicznych roŇõlin. Metoda klonowania in vitro moŇľe byńá stosowana w komercyjnych hodowlach o przeznaczeniu farmaceutycznym.
1 Doyle E, Spence A A. Cannabis as a medicine?. Br J Anaesth 1995; 74: 359-361 2 Hammond C T, Mahlberg P G. Morphogenesis of capitate glandular hairs of Cannabis sativa(Cannabaceae). Am J Bot 1977; 64: 1023-1031 3 Mechoulam R, Ben-Shabat A. From gan-zi-gun-nu to anandamide and 2- arachidonoylglycerol: the ongoing story of Cannabis. Nat Prod Rep 1999; 16: 131-143 4 Sirikantaramas S, Taura F, Morimoto S, Shoyama Y. Recent advances in Cannabis sativa research: biosynthetic studies and its potential in biotechnology. Curr Pharm Biotechnol 2007; 8: 237-243 5 Brenneisen R, Egli A, ElSohly M A, Henn V, Spiess Y. The effect of orally and rectally administered őĒ9-tetrahydrocannabinol on spasticity: a pilot study with 2 patients. Int J Clin Pharmacol Ther 1996; 34: 446-452 6 Formukong E A, Evans A T, Evans F. The medicinal uses of Cannabis and its constitutents. Phytother Res 1989; 3: 219-231 7 Grinspoon L, Bakalar J B. Marihuana, the forbidden medicine. New Haven; Yale University Press 1993 8 Mattes R D, Shaw L M, Eding-Owens J, Egelman K, ElSohly M A. By passing the first pass effect for therapeutic use of cannabinoids. Pharmacol Biochem Behav 1993; 44: 745-747 9 Mattes R D, Egelman K, Shaw L M, ElSohly M A. Cannabinoids appetite stimulation. Pharmacol Biochem Behav 1994; 49: 187-195 10 Mechoulam R. The pharmacohistory of Cannabis sativa. Mechoulam R Cannabinoids as therapeutic agents. Boca Raton, Florida; CRC Press 1986: 1-19 11 ElSohly M A, Ross S A, Mehmedic Z, Arafat R, Yi B, Banahan B F. Potency trends of őĒ9-THC and other cannabinoids in confiscated marijuana from 1980-1997. J Forensic Sci 2000; 45: 24-30 12 Mehmedic Z, Chandra S, Slade D, Denham H, Foster S, Patel A S, Ross S A, Khan I A, ElSohly M A.Potency trends of őĒ9-THC and other cannabinoids in confiscated Cannabis preparations from 1993-2008. J Forensic Sci 2010; , accepted for publication 13 Loh W H T, Hartsel S C, Robertson W. Tissue culture of Cannabis sativa L. and in vitro biotransformation of phenolics. Z Pflanzenphysiol 1983; 111: 395-400 14 Richez-Dumanois C, Braut-Boucher F, Cosson L, Paris M. Multiplication vegetative in vitro du chanvre (Cannabis sativa L.) application a la conservation des clones selections. Agronomie 1986; 6: 487-495 15 Mandolino G, Ranalli P. Advances in biotechnological approaches for hemp breeding and industry. Ranalli P Advances in hemp research. New York; Haworth Press 1999: 185-208 16 Slusarkiewicz-Jarzina A, Ponitka A, Kaczmarek Z. Influence of cultivar, explant source and plant growth regulator on callus induction and plant regeneration of Cannabis sativa L. Acta Biol Craco Series Bot 2005; 47: 145-151 17 Bing X, Ning L, Jinfeng T, Nan G. Rapid tissue culture method of Cannabis sativa for industrial uses. CN Patent 1887043 A 20070103. 2007 18 Lata H, Chandra S, Khan I, ElSohly M A. Propagation through alginate encapsulation of axillary buds ofCannabis sativa L. - an important medicinal plant. Physiol Mol Biol 2009; 15: 79-86 19 Lata H, Chandra S, Khan I, ElSohly M A. Thidiazuron induced high frequency direct shoot organogenesis of Cannabis sativa L. In Vitro Cell Dev Biol Plant 2009; 45: 12-19 20 Ross S A, Parker M, Arafat R, Lovett K, ElSohly M A. The analysis of confiscated marijuana samples for different cannabinoids using GC/FID. Am Lab 1996; 16: 16-17 21 Pierik R L M. Commercial aspects of micropropagation. Prakash J, Pierik RLM Horticulture - new technologies and applications. Dordrecht; Kluywer Academic Publisher 1991: 141-153 22 Shenoy V B, Vasil I K. Biochemical and molecular analysis of plants derived from embryogenic cultures of nipper grass (Pennisetum purpureum K. Schum). Theor Appl Genet 1992; 83: 940-955 23 Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 1962; 15: 473-497 24 Rani V, Parida P, Raina S N. Random amplified polymorphic DNA (RAPD) markers for genetic analysis in micropropagated plants of Populus deltoides Marsh. Plant Cell Rep 1995; 14: 459-462 25 Wang X R, Szmidt A E, Nguyen H N. The phylogenetic position of the endemic flat-needle pine Pinus krempfii (Lec., Pinaceae) from Vietnam, based on PCR-RFLP analysis of chloroplast DNA. Plant Syst Evol 2000; 220: 21-36 26 Damasco O P, Graham G C, Henry R J, Adkins S W, Smith M K, Godwin I D. Random amplified polymorphic DNA (RAPD) detection of dwarf off types in micropropagated Cavendish (Musa spp. AAA) bananas. Plant Cell Rep 1996; 16: 118-123 27 Salvi N D, George L, Eapen S. Plant regeneration from leaf callus of turmeric and random amplified polymorphic DNA analysis of regenerated plants. Plant Cell Tissue Organ Cult 2001; 66: 113-119 28 Ma X, Gang D R. Metabolic profiling of in vitro micropropagated and conventionally greenhouse grown ginger (Zingiber officinale). Phytochemistry 2006; 2239-2255 29 Lata H, Chandra S, Techen N, Khan I A, ElSohly M A. Assessment of the genetic stability of micropropagated plants of Cannabis sativa by ISSR markers. Planta Med 2009; DOI: 10.1055/s-0029-1185945 , advance online publication 30 Chandra S, Lata H, Khan I A, ElSohly M A. Photosynthetic response of Cannabis sativa L. to variations in photosynthetic photon flux densities, temperature and CO2 conditions. Physiol Mol Biol Plants 2008; 14: 299-306
- uŇľyta metoda klonowania in vitro [17](fragmenty Ňāodyg z bocznymi pńôdami) daje wysokńÖ skutecznoŇõńá, oraz gwarantuje identycznoŇõńá genetycznńÖ (z kt√≥rńÖ jest cieŇľko przy klonowaniu z tkanki przyrannej) - jeŇõli pobierzemy klony (zar√≥wno normalne jak i in vitro) i bńôdziemy je trzymańá w takich samych warunkach jak roŇõlinńô matkńô to uzyskamy identyczne stńôŇľenie kanabinoid√≥w - najlepszy wzrost pńôd√≥w przy 0,5 ¬ĶM TDZ w poŇľywce - najlepsze ukorzenianie w poŇľywce zawierajńÖcej 0,25 ¬ĶM TDZ, 500mg/l wńôgla aktywowanego, oraz 2,5 ¬ĶM IBA -stńôŇľenia THC, THCV, CBG, CBD rosnńÖ od fazy wegetatywnej do szczytu kwitnienia, potem spadajńÖ - przy okoŇāo 430W/m2 (HPS i MH) nie uzyskano maksymalnego moŇľliwego stńôŇľenia THC (por√≥wnujńÖc do outdoor'u w Missisipi )
Po raz pierwszy uŇľyto Marihuany w Chinach w roku 1737 jako Lekarstwo ŇāagodzńÖce b√≥l reumatyzmu i artretyzmu.
Pierwsze przepisy, kt√≥re dotyczyŇāy Marihuany nakazywaŇāy farmerom jej uprawńô. UŇľywano Konopi do produkcji ubraŇĄ, Ňľagli i lin.
George Washington i Thomas Jefferson posiadali plantacjńô Konopi.
Jefferson napisaŇā Deklaracjńô NiepodlegŇāoŇõci na papierze z Konopi.
W Ameryce, Marihuana jest czwartńÖ uprawianńÖ roŇõlinńÖ pod wzglńôdem og√≥lnej wartoŇõci i tworzy rynek warty 36 miliard√≥w $.
DziaŇāanie psychoaktywne majńÖ tylko kwiatostany ŇľeŇĄskich roŇõlin. Mńôskie sŇāuŇľńÖ tylko do zapylania ŇľeŇĄskich.
Nazwa Marihuana pochodzi od nieznanego meksykaŇĄskiego etymologa. ZastńÖpiŇāa "Cannabis" i "Konopie" w latach 30.
Rekordowy joint zawiera w sobie 100 gram zioŇāa.
